Single Molecule Enzymatic Assays
Like enzymology in general, single-molecule enzymology has been limited and facilitated by assay developments. Recent advances in single-molecule enzymatic assays have profoundly changed how biochemical reactions are studied, revealing new insights into established problems and opening new lines of investigation. To that end, our group has developed single molecule fluorescence and flow stretching enzymatic assays.
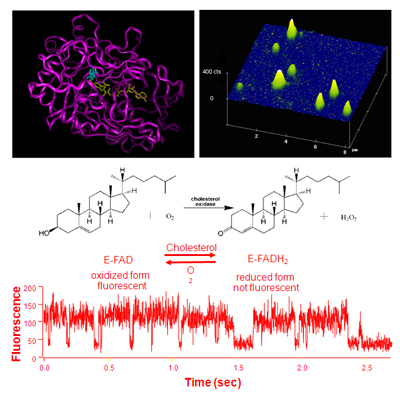
Single Molecule Fluorescence Assays by Fluorescent Measurements
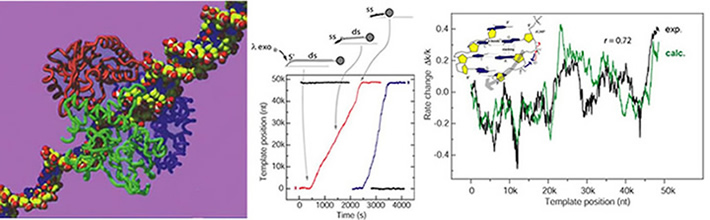
Some of the individual λ exonuclease enzyme molecules digest processively all 48knt of the λ phage DNA substrate. The digestion rate of the individual enzymes exhibits large fluctuations. These fluctuations are partially dependent on the sequence of the λ DNA. We calculate the variations in the enzymatic rate caused by the modulation on the free energy required to melt the cleaved bases (green trace*) and compare it to the average of multiple single-molecule rate traces (black trace). The high degree of correlation identifies the base melting as a rate-limiting step in the catalytic cycle.
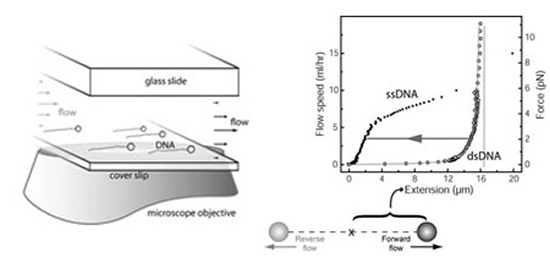
We note that many single molecule assays are based on optical tweezers and magnetic tweezers on nucleic acid enzymes and molecule motors, which were pioneered by the David Bensimon/Vincent Croquette [18], Steve Block [19], Carlos Bustamante [17], and Toshio Yanagida [2] groups.
Single-Molecule Transcription Assay
We have developed a high-throughput in vitro single-molecule assay to monitor repetitive stochastic transcription events in real time on individual DNA templates [20]. We use a nucleic acid stain, SYTO RNASelect, that is nonfluorescent at 530 nm but becomes fluorescent upon binding to RNA. It has been used to detect RNA in the presence of DNA [21]. An Argon laser line at 488 nm is used to excite SYTO RNASelect in a total internal reflection fluorescence (TIRF) microscope. We collect fluorescence at 530 nm and record time-lapse movies with a CCD camera. In the presence of the dye, a single nascent mRNA becomes visible, and its fluorescence intensity increases with the mRNA length.
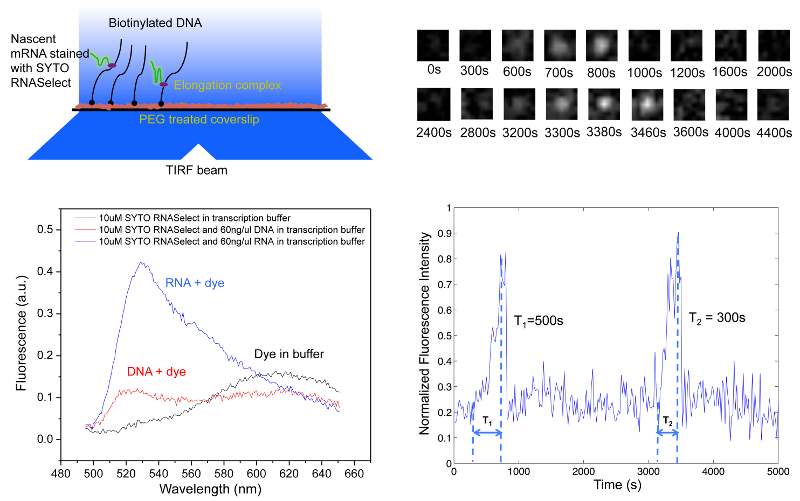
We tether DNA templates containing a promoter on the passivated surface of the flow cell through biotin-streptavidin linkage. After we infuse a mixture of RNA polymerase, NTPs and SYTO RNASelect into the flow cell, the fluorescence intensity of many spots in the field of view linearly ramps up due to transcription elongation, which is followed by abrupt disappearance upon transcription termination. “Blinking” of fluorescence occurs when multiple transcripts are produced from a template. By recording fluorescent movies, we could monitor transcription elongation in real time as nascent mRNAs are produced on hundreds of templates simultaneously.
Transcription is known to generate DNA positive supercoiling ahead of the elongation complex and negative supercoiling behind it when the free spiral of the bulky complex around the DNA is hindered by the frictional drag on the complex [22,23]. This single-molecule assay allows us to investigate the effects of supercoiling on transcription initiation and elongation in a clean and controlled system.
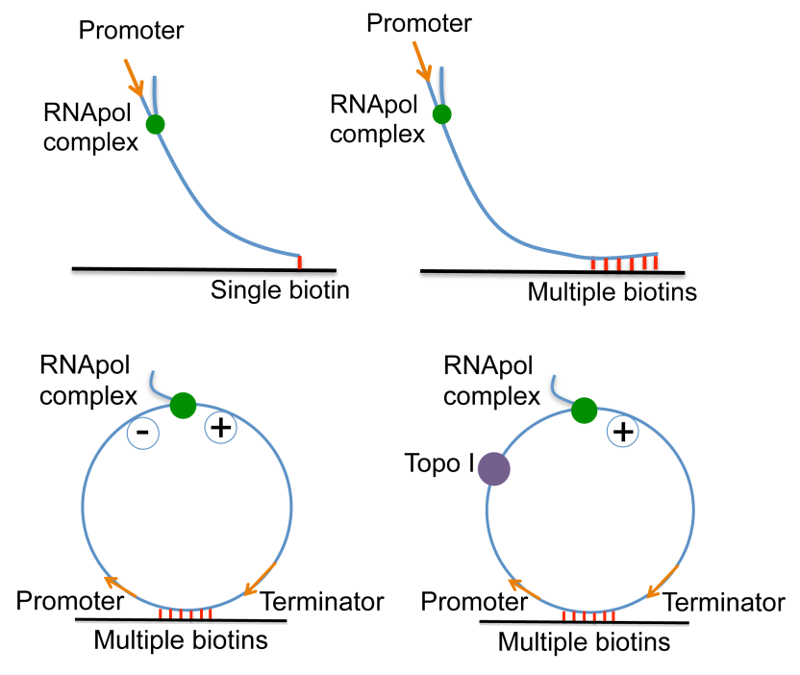
In order to examine the effect of positive supercoiling buildup on transcription elongation, we design 12 kb-long linear DNA templates with a promoter on the 5’ end, and single or multiple biotinylated nucleotides on the 3’ end. On the single-biotin template, supercoiling cannot accumulate since DNA can rotate around its single linkage to the surface. In contrast, on the multiple-biotin template, DNA cannot rotate around its multiple linkages, thus positive supercoiling would accumulate downstream of the elongation complex. We have shown that positive supercoiling generated by transcription slows down elongation [20].
In order to examine the effect of positive supercoiling on transcription initiation, we design a circular template and tether it to the surface with multiple biotin-streptavidin linkages. The circular template consists of a promoter, a 12 kb-long transcribing sequence and a terminator. In the presence of E. coli Topoisomerase I, negative supercoiling generated by transcription is continuously removed, and positive supercoiling accumulates on the circular template as multiple transcripts are made from the template. We have shown that positive supercoiling generated by transcription stops initiation [20].
We note our assay is different from other existing in vitro transcription assays using single-molecule manipulation or single-molecule fluorescence imaging. Our assay uses RNA staining so that the elongation process on templates with any sequence can be easily monitored for multiple rounds of transcription on each template. Meanwhile, this is a high-throughput measurement because hundreds of templates in one field of view can be monitored simultaneously. This assay will be generally useful for studying other questions in transcription, such as pausing and termination kinetics.
References
[1] H.P. Lu, L. Xun, X.S. Xie. Science 282, 1877 (1998).
[2] Funatsu T., et al. Nature 374, 555 (1995).
[3] M.J. Levene, J. Korlach, S.W. Turner, M. Foquet, H.G. Graighead, W.W. Webb. Science 299, 682–686 (2003).
[4] T. Ha, et al. PNAS 96, 893–898 (1999).
[5] X. Zhuang, et al. Science 288, 2048–2051 (2000).
[6] C. Yu, D. Hu, E.R. Vorpagel, H.P. Lu. J. Phys. Chem. B. 107, 7947 (2003).
[7] R. Yasuda, H. Noji, K. Kinosita, M.Yoshida. Cell, 93, 1117 (1998).
[8] A. Yildiz, J. N. Forkey, S. A. McKinney, T. Ha, Y. E. Goldman, P.R. Selvin. Science 300, 2016, (2003).
[9] G. Luo, et al., PNAS 104, 12610–12615 (2007).
[10] L. Edman et al. Chem. Phys. 247, 11–22 (1999).
[11] B. English, et al. Nat. Chem. Biol. 2, 87–94 (2006).
[12] K. Velonia, O. Flomenbom, D. Loos, S. Masuo, M. Cotlet, Y. Engelborghs, J. Hofkens, A.E. Rowan, J. Klafter, R.J.M. Nolte, F.C. de Schryver. Angew. Chem. Int. Ed. (2004) 43, 2–6 (2005).
[13] A.M. van Oijen, P.C. Blainey, D.J. Crampton, C.C. Richardson, T. Ellenberger, X.S. Xie. Science 301, 1235 (2003).
[14] C.M. Schroeder, P.C. Blainey, S. Kim, X.S. Xie. In “Single Molecule Techniques A Laboratory Manual” edited by P.R. Selvin and T. Ha, p 461-492, Cold Spring Harbor Laboratory Press, 2008.
[15] S. Kim, P.C. Blainey, C.M. Schroeder, X.S. Xie. Nature Methods, 4, 397-399 (2007).
[16] S.B. Smith, L. Finzi, C. Bustamante. Science 258, 1122 (1992).
[17] G.J.L. Wuite, S.B. Smith, M. Young, D. Keller, C. Bustamante. Nature 404, 103 (2000).
[18] B. Maier, D. Bensimon, V. Croquette. PNAS 97, 12002 (2000).
[19] K. Svoboda, C.F. Schmidt, B.J. Schnapp, S.M. Block. Nature, 365, 721 (1993).
[20] S. Chong, C. Chen, H. Ge, X.S. Xie. Cell 158, 314-326 (2014).
[21] C. Kannemeier et al. PNAS 104, 6388-6393 (2007).
[22] L.F. Liu, J.C. Wang. PNAS 84, 7024-7027 (1987).
[23] Y.P. Tsao, H.Y. Wu, L.F. Liu. Cell 56, 111-118 (1989).